

European Journal of Neurodegenerative Diseases 2024; 13(1) January-April: 23-28
THE ROLE OF SEROTONIN IN IMMUNITY AND INFLAMMATION
F. Gaudelli 1* and C. Annicchiarico 2
1 School of Dentistry, University of Foggia, Foggia, Italy;
2 Independent researcher, 70100 Bari, Italy.
*Correspondence to:
Federico Gaudelli, DDS
School of Dentistry,
University of Foggia,
71100 Foggia, Italy.
e-mail: federico_gaudelli.563780@unifg.it
| Received: 09 February, 2024 Accepted: 20 February, 2024 |
2974-6345 (2024) Copyright © by BIOLIFE This publication and/or article is for individual use only and may not be further reproduced without written permission from the copyright holder. Unauthorized reproduction may result in financial and other penalties. Disclosure: all authors report no conflicts of interest relevant to this article. |
ABSTRACT
Serotonin (5-HT) is a neurotransmitter that regulates different functions of the human body. 5-HT has immunomodulatory effects and plays an important role in the brain, regulating body temperature, mood, emotions, sexuality, sleep, appetite, and cognitive functions, amongst other functions. In the immune system, this neurotransmitter is produced by monocytes/macrophages, lymphocytes, and mast cells (MCs), and interacts with circulating immune cells in peripheral tissues. 5-HT production can be enhanced by increases of cytokines such as IL-1β, IL-33, and IL-13, and can have both inhibitory and stimulatory effects on innate immune cells. At physiological levels, 5-HT can stimulate the immune system, which can be helpful for the body, but at elevated concentrations, it can activate inflammation and be harmful.
KEYWORDS: serotonin, 5-HT, neurotransmitter, immunity, inflammation, CNS
INTRODUCTION
Neurotransmitters are proteins that transmit information across synapses between neurons. They are contained in the synaptic vesicles of neurons that fuse to the neuronal membrane after a stimulus and release neurotransmitters into the synaptic cleft. These neuropeptides bind to their specific receptors and/or ion channels located on the postsynaptic neuron membrane and cause a biological response. Neurotransmitters can be derived from amino acids such as aspartic acid, glutamic acid, glycine, and γ-aminobutyric acid (GABA) (Table 1).
Table I. Some of the most well-known neurotransmitters.
| Monoamine (from phenylalanine and tyrosine): | dopamine, norepinephrine, epinephrine |
| From tryptophan: | serotonin (5-HT), melatonin, histidine, histamine |
| Peptides and neuropeptides: | neurotensin (NT), galanin, bombesin, gastrin-releasing peptide (GRP), neuromedin B |
| neurohypophysis: | vasopressin, oxytocin, neurophysin (type I and II), neuropeptide Y, pancreatic polypeptide, peptide YY |
| Tachykinins: | neurokinin A, neurokinin B, neuropeptide A, gamma neuropeptide, substance P |
| Other neurotransmitters: | acetylcholine, glucagon, vasoactive intestinal peptide (VIP), somatostatin, endorphin, enkephalin |
Serotonin or 5-hydroxy tryptamine (5-HT) (molecular formula: C10H12N2O) was discovered in 1935, when it was isolated in Pavia by Vittorio Erspamer (1) (Fig.1). 5-HT is a neurotransmitter that plays an important role in the brain. 5-HT is synthesized from tryptophan, an amino acid produced in high quantities by the neurons of the central nervous system (CNS) and in the enterochromaffin cells of the gastrointestinal system (2). Levels of 5-HT are more concentrated in different areas of the brain; For example, the hypothalamus contains more 5-HT than the neopallium and cerebellum, where the quantity is lower. 5-HT levels depend on the amount of dietary tryptophan that enters the CNS (3).

Fig. 1. The chemical structure of serotonin (5-HT).
The precursor amino acid of 5-HT is tryptophan, which is converted to 5-hydroxytryptophan (5-HTP) from hydroxylated tryptophan. Subsequently, the 5-HTP is decarboxylated by the enzyme decarboxylase to form 5-HT (4) (Fig.2). 5-HT is stored in neurons as a neurotransmitter and cooperates with hormones including substance P and somatostatin. The degradation of 5-HT is carried out by the monoamine oxidase (MAO) pathway which produces 5-hydroxyindole acetic acid (5-HIAA), and by the sulfotransferase enzyme that leads to the generation of tryptamine-O-sulphate with a reduction of 5-HT.
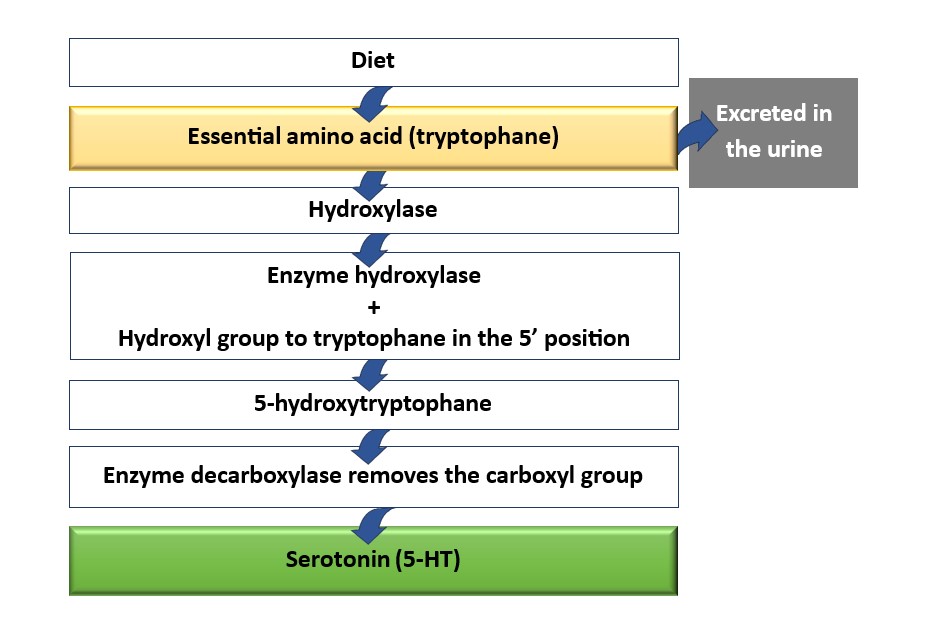
Fig. 2. The biochemical cascade for serotonin (5-HT) synthesis, starting from tryptophan.
5-HT is dispersed differently throughout the body, with some organs having a higher concentration. The walls of the small intestines and stomach possess enterochromaffin cells with high amounts of 5-HT, which regulates intestinal motility and secretion (5). Platelets are another source of 5-HT, which is released into the bloodstream, acts on the dilation of vessels, and helps regulate homeostasis and heal wounds (6). 5-HT is produced by nine brain nuclei In the CNS that extend to the spinal cord, and it performs various functions such as regulation of body temperature, mood, emotions, sexuality, cognitive functions, sleep, creativity, and appetite (7,8) (Table II). In mice, 5-HT causes pain if injected subcutaneously (9). Additionally, 5-HT plays an important role in the immune system where it is involved in interactions with T cells, mast cells (MCs), macrophages, dendritic cells, and platelets (10).
Table II. Some functions of the human body that are regulated by serotonin.
| · Mood and emotions | · Sexual health | · Nausea |
| · Sleep | · Bone health | · Cognitive functions |
| · Digestion and appetite | · Wound healing | · Body temperature |
The role of serotonin (5-HT) in immunity and inflammation
5-HT has immunomodulatory effects and interacts with circulating immune cells in peripheral tissues (11). It seems that cells such as monocytes/macrophages, lymphocytes, MCs, platelets, vascular smooth muscle cells, and adipocytes are all sources of 5-HT in the human body (12). However, 5-HT is mostly stored in and released by platelets after activation (13).
5-HT that is synthesized in the brain and peripheral tissues causes various effects and represents a classic neurotransmitter with both autocrine and paracrine action. 5-HT is implicated in peristaltic motility and gastric disorders, and in intestinal secretion (14). 5-HT is involved in the immune response and is released by endothelial cells and then subsequently stored in platelet granules, which are a source of 5-HT for the cells of the immune system. 5-HT participates in innate and adaptive immunity after being secreted by macrophages, MCs, and T lymphocytes (15).
In humans, there are 7 types of 5-HTR receptors, which are numbered from 1 to 7 (5-HTR1-5-HTR7) and are involved in different functions. For example, the 5-HTR1 and 5-HTR5 receptors downregulate the synthesis of cyclic adenosine monophosphate (cAMP) (16), and 5-HTR4, 6, and 7 activate adenylate cyclase and promote cAMP activity (17). The activation of 5-HTR2 stimulates inositol triphosphate and diacylglycerol and increases Ca2+ fluxes in the cell cytoplasm, and 5-HTR3 causes the influx of Ca2+ and Na+ with the exit of K+, causing a depolarization of the cell membrane (18). In turn, 5-HTR1 includes 5 subtypes ranging from A to F (but not including C), 5-HTR2 has 3 subtypes (A, B, and C), while 5-HTR3, 4, 5, 6, and 7 have no subtypes.
5-HT that is produced by endothelial cells can exert an effect on circulating naïve lymphocytes, influencing the immune system (10). An increase in the cytokines IL-1β, IL-33, and IL-13 has been reported to enhance 5-HT production from endothelial cells (19)
In animal experiments, it was noted that injecting 5-HT into the cerebral ventricles sharply elevated body temperature (probably by mediating IL-1 activation), while catecholamines lowered it, actions that are mediated by the hypothalamus (20). 5-HT can have both stimulatory and inhibitory effects on innate immune cells, such as macrophages, depending on the dose administered. With low concentrations of interferon gamma (IFN-g) stimulation, 5-HT can increase phagocytosis, while with high concentrations of IFN-g, 5-HT inhibits phagocytosis (21). In in vitro studies, macrophages treated with 5-HT binding the 5-HTR1A receptor secrete more pro-inflammatory cytokines than untreated macrophages (19), an effect that involves the upregulation of nuclear factor kappa B (NF-kB) activation. In addition, 5-HT stimulates the production of tumor necrosis factor (TNF) and upregulates the expression of M2 polarization through the activation of 5-HTR2B and 5-HTR7 (22).
By activating the 5-HTD1 receptors, 5-HT contracts blood vessels, including those in the brain which, when dilated, cause migraines (23). In addition, by binding to 5-HT3 receptors, 5-HT stimulates nociceptive sensory nerve endings and plays a key role in the excitation and inhibition of neurons and in the regulation of CNS functions (24). 5-HT elevates cortisol and prolactin levels, inhibiting gonadotropin-releasing factor and inhibiting ejaculation (25). In fact, low levels of 5-HT cause premature ejaculation (26). 5-HT dysfunction has been associated with neuropsychiatric disorders including bulimia and anorexia (27), anxiety (28), schizophrenia (29), and obsessive-compulsive disorder (30), although the mechanisms by which 5-HT causes these diseases is not yet clear, and it could be that targeting 5-HT may represent new therapeutic strategies (Table III).
Table III. Neurological effects associated with low serotonin (5-HT) levels.
| · Anxiety | · Obsessive-compulsive disorder |
| · Sleep problems | · Schizophrenia |
| · Depression | · Suicidal behavior |
| · Panic disorders | · Phobias |
| · Bulimia | · Post-traumatic stress disorder |
Sometimes 5-HT deficiency can provoke depression; in fact, it is called the “happiness hormone”. The effects of 5-HT on depression were found to be conflicting in different studies (31). When administered through the diet, tryptophan, the essential amino acid precursor of serotonin, was seen to alleviate behaviors associated with depression and anxiety in mice (32). It was seen that the reduction of tryptophan introduced through the diet was not able to induce depression, although animal studies may not reflect those on man.
Tryptophan is an essential aromatic amino acid and its metabolism is mediated by intestinal bacterial degradation that occurs through two pathways: 5-HT bioamine and kynurenine (19). Tryptophan metabolites are involved in the immune response and in brain functioning. Studies have reported that pharmacologically inhibiting tryptophan hydroxylase can cause depression in around 30% of cases. (33,34).
The binding of 5-HT on the 5-HTR1A and 5-HTR2A receptors of natural killer (NK) cells has a stimulatory effect with increased cytotoxicity and IFN-g secretion (35). 5-HT also has an anti-apoptosis action and an action against oxidative damage to NK cells. Furthermore, in depression patients treated with 5-HT inhibitors, increased cytotoxicity and proliferation of NK cells has been noted (36). All these effects strictly depend on the concentration of 5-HT used, which can give opposite results.
CONCLUSIONS
Therefore, in conclusion, the concentration of 5-HT seems to affect the actions of the immune system, even if the data in the literature appears to be conflicting. At physiological concentrations, 5-HT can stimulate the immune system, while at high concentrations, it can activate inflammation and be harmful to health.
Conflict of interest
The authors declare that they have no conflict of interest.
REFERENCES
- Erspamer V. Pharmacology of enteramine. I. Action of acetone extract of rabbit stomach mucosa on blood pressure and on surviving isolated organs. Archiv fur Experimentelle Pathologie und Pharmakologie. 1940;196:343-346.
- Gershon MD. 5-Hydroxytryptamine (serotonin) in the gastrointestinal tract. Current Opinion in Endocrinology & Diabetes and Obesity. 2013;20(1):14-21. doi:https://doi.org/10.1097/med.0b013e32835bc703
- Höglund E, Øverli Ø, Winberg S. Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Frontiers in Endocrinology. 2019;10. doi:https://doi.org/10.3389/fendo.2019.00158
- Maffei ME. 5-Hydroxytryptophan (5-HTP): Natural Occurrence, Analysis, Biosynthesis, Biotechnology, Physiology and Toxicology. International Journal of Molecular Sciences. 2020;22(1):181. doi:https://doi.org/10.3390/ijms22010181
- Liu N, Sun S, Wang P, Sun Y, Hu Q, Wang X. The Mechanism of Secretion and Metabolism of Gut-Derived 5-Hydroxytryptamine. International Journal of Molecular Sciences. 2021;22(15):7931. doi:https://doi.org/10.3390/ijms22157931
- Machida T, Iizuka K, Hirafuji M. 5-hydroxytryptamine and its receptors in systemic vascular walls. Biological & Pharmaceutical Bulletin. 2013;36(9):1416-1419. doi:https://doi.org/10.1248/bpb.b13-00344
- Olivier B. Serotonin: A never-ending story. European Journal of Pharmacology. 2015;753:2-18. doi:https://doi.org/10.1016/j.ejphar.2014.10.031
- Ciranna L. Serotonin as a Modulator of Glutamate- and GABA-Mediated Neurotransmission: Implications in Physiological Functions and in Pathology. Current Neuropharmacology. 2006;4(2):101-114. doi:https://doi.org/10.2174/157015906776359540
- Sufka KJ, Schomburg FM, Giordano J. Receptor mediation of 5-HT-induced inflammation and nociception in rats. Pharmacology Biochemistry and Behavior. 1992;41(1):53-56. doi:https://doi.org/10.1016/0091-3057(92)90058-n
- Ahern GP. 5-HT and the immune system. Current Opinion in Pharmacology. 2011;11(1):29-33. doi:https://doi.org/10.1016/j.coph.2011.02.004
- Roszman TL, Jackson JC, Cross RJ, Titus MJ, Markesbery WR, Brooks WH. Neuroanatomic and neurotransmitter influences on immune function. The Journal of Immunology. 1985;135(2):769-772. doi:https://doi.org/10.4049/jimmunol.135.2.769
- Herr N, Bode C, Duerschmied D. The Effects of Serotonin in Immune Cells. Frontiers in Cardiovascular Medicine. 2017;4. doi:https://doi.org/10.3389/fcvm.2017.00048
- McNicol A, Israels SJ. Platelet Dense Granules: Structure, Function and Implications for Haemostasis. Thrombosis Research. 1999;95(1):1-18. doi:https://doi.org/10.1016/S0049-3848(99)00015-8
- Mawe GM, Hoffman JM. Serotonin signalling in the gut—functions, dysfunctions and therapeutic targets. Nature Reviews Gastroenterology & Hepatology. 2013;10(8):473-486. doi:https://doi.org/10.1038/nrgastro.2013.105
- Hodo TW, de Aquino MTP, Shimamoto A, Shanker A. Critical Neurotransmitters in the Neuroimmune Network. Frontiers in Immunology. 2020;11. doi:https://doi.org/10.3389/fimmu.2020.01869
- Canesi L, Miglioli A, Balbi T, Fabbri E. Physiological Roles of Serotonin in Bivalves: Possible Interference by Environmental Chemicals Resulting in Neuroendocrine Disruption. Frontiers in Endocrinology. 2022;13:792589. doi:https://doi.org/10.3389/fendo.2022.792589
- Graveleau C, Paust HJ, Schmidt-Grimminger D, Mukhopadhyay AK. Presence of a 5-HT7Receptor Positively Coupled to Adenylate Cyclase Activation in Human Granulosa-Lutein Cells1. The Journal of Clinical Endocrinology & Metabolism. 2000;85(3):1277-1286. doi:https://doi.org/10.1210/jcem.85.3.6448
- Rondé P, Nichols RA. High calcium permeability of serotonin 5-HT3 receptors on presynaptic nerve terminals from rat striatum. Journal of Neurochemistry. 1998;70(3):1094-1103. doi:https://doi.org/10.1046/j.1471-4159.1998.70031094.x
- Haq S, Grondin JA, Khan WI. Tryptophan‐derived serotonin‐kynurenine balance in immune activation and intestinal inflammation. The FASEB Journal. 2021;35(10). doi:https://doi.org/10.1096/fj.202100702r
- Feldberg W, Myers RD. Temperature changes produced by amines injected into the cerebral ventricles during anaesthesia. The Journal of Physiology. 1964;175(3):464-478. doi:https://doi.org/10.1113/jphysiol.1964.sp007527
- Freire-Garabal M, Núñez MJ, Balboa J, et al. Serotonin upregulates the activity of phagocytosis through 5-HT1A receptors. British Journal of Pharmacology. 2003;139(2):457-463. doi:https://doi.org/10.1038/sj.bjp.0705188
- Domínguez-Soto Á, Usategui A, Casas-Engel ML, et al. Serotonin drives the acquisition of a profibrotic and anti-inflammatory gene profile through the 5-HT7R-PKA signaling axis. Scientific Reports. 2017;7(1):14761. doi:https://doi.org/10.1038/s41598-017-15348-y
- Tepper SJ, Rapoport AM, Sheftell FD. Mechanisms of Action of the 5-HT1B/1D Receptor Agonists. Archives of Neurology. 2002;59(7):1084. doi:https://doi.org/10.1001/archneur.59.7.1084
- Pithadia AB, Jain SM. 5-Hydroxytryptamine Receptor Subtypes and their Modulators with Therapeutic Potentials. Journal of Clinical Medicine Research. 2009;1(2). doi:https://doi.org/10.4021/jocmr2009.05.1237
- Freeman ME, Kanyicska B, Lerant A, Nagy G. Prolactin: structure, function, and regulation of secretion. Physiological reviews. 2000;80(4):1523-1631. doi:https://doi.org/10.1152/physrev.2000.80.4.1523
- Giuliano F, Clément P. Serotonin and Premature Ejaculation: From Physiology to Patient Management. European Urology. 2006;50(3):454-466. doi:https://doi.org/10.1016/j.eururo.2006.05.055
- Bailer UF, Kaye WH. Serotonin: imaging findings in eating disorders. Current Topics in Behavioral Neurosciences. 2011;6:59-79. doi:https://doi.org/10.1007/7854_2010_78
- Garcia-Garcia AL, Newman-Tancredi A, Leonardo ED. P5-HT1A receptors in mood and anxiety: recent insights into autoreceptor versus heteroreceptor function. Psychopharmacology. 2013;231(4):623-636. doi:https://doi.org/10.1007/s00213-013-3389-x
- Akhondzadeh S. The 5-HT hypothesis of schizophrenia. PubMed. 2001;4(3):295-300.
- Murphy DL, Zohar J, Benkelfat C, Pato MT, Pigott TA, Insel TR. Obsessive—Compulsive Disorder as a 5-HT Subsystem-Related Behavioural Disorder. British Journal of Psychiatry. 1989;155(S8):15-24. doi:https://doi.org/10.1192/s0007125000291721
- Graeff FG, Guimarães FS, De Andrade TGCS, Deakin JFW. Role of 5-HT in stress, anxiety, and depression. Pharmacology Biochemistry and Behavior. 1996;54(1):129-141. doi:https://doi.org/10.1016/0091-3057(95)02135-3
- Wang D, Wu J, Zhu P, et al. Tryptophan-rich diet ameliorates chronic unpredictable mild stress induced depression- and anxiety-like behavior in mice: The potential involvement of gut-brain axis. Food Research International. 2022;157:111289. doi:https://doi.org/10.1016/j.foodres.2022.111289
- Ruhé HG, Mason NS, Schene AH. Mood is indirectly related to serotonin, norepinephrine and dopamine levels in humans: a meta-analysis of monoamine depletion studies. Molecular Psychiatry. 2007;12(4):331-359. doi:https://doi.org/10.1038/sj.mp.4001949
- Cowen PJ, Browning M. What has serotonin to do with depression? World Psychiatry. 2015;14(2):158-160. doi:https://doi.org/10.1002/wps.20229
- Kanova M, Kohout P. Serotonin—Its Synthesis and Roles in the Healthy and the Critically Ill. International Journal of Molecular Sciences. 2021;22(9):4837. doi:https://doi.org/10.3390/ijms22094837
- Patas K, Willing A, Demiralay C, et al. T Cell Phenotype and T Cell Receptor Repertoire in Patients with Major Depressive Disorder. Frontiers in Immunology. 2018;9. doi:https://doi.org/10.3389/fimmu.2018.00291