

European Journal of Neurodegenerative Diseases 2023; 12(1) January-April: 5-10
IMPACT OF INSULIN-LIKE GROWTH FACTORS 1 AND 2 IN THE INFLAMMATORY RESPONSE MEDIATED BY CYTOKINES
P. Conti 1,2* and I. Tsilioni 1
1 Laboratory of Molecular Immunopharmacology and Drug Discovery, Tufts University, School of Medicine, Boston, MA, USA;
2 Former Professor of Immunology, Postgraduate Medical School, University of Chieti-Pescara, Italy.
*Correspondence to:
Professor Pio Conti,
Affiliated Professor of Molecular Immunopharmacology and Drug Discovery,
Tufts University School of Medicine,
Boston, MA, USA.
e-mail: pioconti@yahoo.it
| Received: 29 March, 2023 Accepted: 27 April, 2023 |
2974-6345 (2023) Copyright © by BIOLIFE This publication and/or article is for individual use only and may not be further reproduced without written permission from the copyright holder. Unauthorized reproduction may result in financial and other penalties. Disclosure: all authors report no conflicts of interest relevant to this article. |
ABSTRACT
In inflammatory disorders such as pancreatitis, immune cells are activated and produce pro-inflammatory cytokines and chemokines, including IL-1, IL-6, and tumor necrosis factor (TNF). These cytokines mediate the immune response while insulin-like growth factors (IGFs), hormones that promote physiological growth, also participate in the inflammatory response. Macrophages play an important role in this process since polarized M1 macrophages provoke acute inflammation, while the polarized M2 type is involved in the anti-inflammatory response and the development of tissue fibrosis. Neuroendocrine and metabolic responses are also present during the inflammatory process, and there is pituitary generation of IGF. A mitogen-activated protein kinase (MAPK) phosphorylation cascade is activated with the binding of IGF-1 to its receptor IGF-1R, which leads to gene expression and key biological responses such as cell proliferation. An increase of IGF-2 and its receptor IGF-2R is associated with diverse biological effects including hypertrophy and cardiac contraction, increases in cardiomyocytes and cardiomyocyte apoptosis, and decreased muscle regeneration and capillary remodeling, amongst others. IGF-2 also mediates chondrocyte hypertrophy with an increase in oxidative phosphorylation, as the inhibition of this process damages hypertrophic differentiation. In this article, we discuss the impact of IGFs in the inflammatory response that is mediated by cytokines.
KEYWORDS: insulin-like growth factor, inflammation, cytokine, myokine, IGF-1, IGF-2, macrophage, IL-1, TNF
INTRODUCTION
Numerous inflammatory mediators, such as pro-inflammatory cytokines, are involved in pancreatitis where immune cells are activated, including macrophages that produce IL-1, tumor necrosis factor (TNF), and other pro-inflammatory cytokines and chemokines. Macrophages are known to play an important role in the pathogenesis of pancreatitis. They can be polarized as M1, that become protagonists of acute inflammation, while those that are polarized as M2 participate in the anti-inflammatory process and promote the development of pancreatic tissue fibrosis (1).
In addition, activated macrophages, in collaboration with IL-18 and IL-3, can stimulate mast cells (MCs) to produce IL-4 and IL-13 cytokines which increase the number of M2 macrophages (2) (Fig.1).
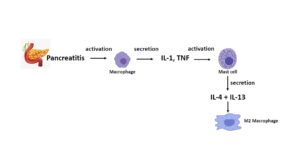
Fig. 1. Diagram depicting the involvement of macrophages and mast cells in the pathogenesis of pancreatitis. Macrophages (M1) become activated and secrete IL-1 and tumor necrosis factor (TNF) amongst other pro-inflammatory cytokines and chemokines. These cytokines activate mast cells (MCs), which secrete IL-4 and IL-13, increasing the amount of M2 macrophages.
Activated M2 macrophages are divided into M2a, M2b, M2c and M2d and are mainly involved in anti-inflammatory responses, while M1 macrophages are involved in pro-inflammatory ones (1). In acute pancreatitis, damaged pancreatic cells release molecules such as zymogen, trypsin, and other cell degradation compounds which activate and recruit macrophages that secrete pro-inflammatory cytokines such as IL-1, TNF, IL-6, interferon-γ (IFNγ), and IL- 18, which participate in the resulting damage (3). The cytokines that are released from the muscle are called myokines and regulate myogenesis and muscle hypertrophy. Myokines can have a systemic action on the immune system and on the adipose tissue that accumulates in the viscera following physical inactivity. The accumulation of adipose tissue results in low-grade inflammation mediated by the recruitment of monokine-releasing macrophages including TNF, IL-1, and IL-6 (4). On the other hand, physical activity activates muscles to produce myokines, such as the above cytokines, which participate in muscle growth and hypertrophy. Physical activity increases insulin sensitivity and fat oxidation leading to the reduction of the inflammatory state.
Insulin growth factor binding proteins (IGFBPs) bind insulin-like growth factor (IGF)-1 and IGF-2 with high affinity, causing biological effects (5). IGF is ubiquitous in all human and rodent fetal tissues and tends to decrease after birth, although, in humans, the serum and tissue levels of IGF-2 remain elevated (6).
IGF-1
IGF-1 is a growth factor of human muscles and bones that is synthesized in the liver and carries out its biological activity by binding to the IGF-1R receptor. IGF-1 mediates bone homeostasis and activates the mitogen-activated protein kinase (MAPK) pathway resulting in cell proliferation (7). MAPK is important in signal transduction and phosphorylation, crucial biological activities for cell signaling (8) (Fig.2).

Fig. 2. Insulin growth factor (IGF)-1 binds to the IGF-R1 receptor, which results in the activation of the mitogen-activated protein kinase (MAPK) pathway with protein kinase cascades; activation of a MAPK kinase kinase (MAPKKK) phosphorylates and activates the MAPKK, which, in turn, activates the MAPK that phosphorylates different substrates in the cytosol and nucleus, resulting in changes in gene expression that cause the biological response of cell proliferation.
IGF-1 contributes to maintaining bone homeostasis and induces osteogenesis by acting on osteoclasts through receptor activator of nuclear factor kappa beta (RANKL) in in vitro experiments (9). In some experiments it has been observed that the overexposed IGF-1 in the osteoblasts leads to an elongation of the bones, while its inhibition reduces bone volume (10). It has been reported that IGF-1 is important for cell development such as chondrocytes and bone growth and plays a crucial role in metabolism by regulating energy function by acting on mitochondria (11). This effect would increase cell proliferation and differentiation, as well as protein synthesis (10). It appears that IGF-1 is instrumental in osteoblast differentiation since the lack of the IGF-1R receptor inhibits bone development. Therefore, IGF-1 is important for the correct development of the skeleton and is correlated with increased levels of this myokine in the muscle where it participates in its growth. In fact, a decrease of IGF-1 in muscle, where it is abundant, can lead to a reduction in bone development (12). Therefore, IGF-1 can be of ubiquitous circulating origin, and is particularly present in bone and muscle, playing a crucial role in bone metabolism.
IGF-2
IGF-2 is an endocrine hormone included in the family of three hormones that possess an insulin-like structure. In humans, the IGF-2 gene is located on chromosome 11 at position p15.5, while in mice, the gene is located on chromosome 7. This peptide has biological activity similar to insulin, with mitogenic and cell growth regulating activity (13). IGF-2 is part of the IGF family made up of IGF-1 and IGF-2, whose biological activity is regulated by 6 proteins ranging from IGFBP-1 to 6 (14).
IGF-2 is the ubiquitous ligand of the IGF-2R receptor and has a crucial role in the physiological development and differentiation of the mouse embryo mesoderm (15). In rodents, IGF-2 is a highly expressed embryonic growth factor, already present in the first days of embryonic development, while it is decreased after birth (16, 17). The binding of IGF-2 to its receptor IGF-2R could favor both tumor growth and onset and influence proliferative cellular pathologies (18). In various inflammatory diseases, such as muscle, rheumatic and cardiac diseases, it has been observed that the expression of IGF-2R is increased, favoring these pathological phenomena (19). Therefore, targeting the IGF-2R receptor could be a new therapeutic approach for proliferative cellular diseases.
The increase of IGF-2 has an important positive action on the development of apoptosis, a phenomenon that occurs in tumors and where IGF-2 and its receptor IGF-2R could have a tumor suppressor effect. Lack of expression of the IGF-2R gene delays the onset of breast cancer, but also the onset of other cancers, an effect that could be inconsistent with the activity of IGF-2 in cervical tumors or glioblastoma, where it acts as an oncogene (20). Inflammation and fibrosis can occur in dystrophic muscle with consequent overexpression of IGF-2 with action on fibroblasts, pericyte endothelial cells, myocytes, and cardiomyocytes (21, 22). In the repair process, immune cells such as macrophages, the complement system, MCs, lymphocytes and neutrophils are involved. The recruitment of these cells regulates the activation, proliferation, and differentiation of muscle cells (Table I).
Table I. Some biological effects caused by an increase of IGF-2 after binding its receptor IGF-2R.
| Increase of: | · Hypertrophy and cardiac contraction |
| · Cardiomyocyte apoptosis | |
| · Size of cardiomyocytes | |
| · EPC migration, adhesion, and invasion | |
| · Elasticity and adhesion of the vascular SHC | |
| · Development of tumors | |
| Decrease of: | · Muscle regeneration |
| · Capillary remodeling | |
| · Muscle force |
IGF-2R performs an important function in suppressing IGF-1 signaling, an effect that could be useful against tumor development (23).
Compared to IGF-1, IGF-2 appears to be more effective against apoptosis (or programmed cell death) in the cells of the placenta. IGF-1 promotes the growth of muscle, which is a major storehouse of energy molecules required for gluconeogenesis. IGF-1 is an important hormone for the energy system and the immune system, regulating the synthesis of some minerals (24). The factors responsible for the storage of energy-rich fuels include insulin IGF-1, testosterone, estrogens, vitamin D, and others, while the factors involved in their release include the renin-angiotensin-aldosterone system, the sympathetic nervous system, the hypothalamic-pituitary-thyroid axis, and the hypothalamic-pituitary-adrenal axis (25).
Somatotropin regulates IGF-2 during intrauterine life, promoting growth during gestation, in contrast to IGF-1, which is synthesized later in adult life. IGF-2 carries out its biological activity through binding to its receptor IGF-1R. During the maturation process of the follicles, the connective cells synthesize and secrete IGF-2, promoting follicular proliferation together with the hormone follicular stimulating hormone (FSH) (26). Subsequently, after ovulation, IGF-2 stimulates the secretion of the hormone progesterone which acts synergistically with the luteinizing hormone (LH) (27). IGF-2 has also been reported to promote survival of hippocampal neurons in newborns, a finding that may have therapeutic implications (28).
By acting on mitochondria, IGF-2 has anti-apoptotic properties, inhibiting cell death and improving the survival of cells such as islet transplantation and cartilage cells (29). It has been reported that physical activity allows for important physiological development through the muscle production of cytokines (also called myokines), including IGFs that perform an anti-inflammatory action (30).
The inflammatory process and the role of pro-inflammatory cytokines
During the inflammatory process, there are neuroendocrine and metabolic responses, and pituitary generation of IGF, which allows for cell survival. Cytokines have autocrine, paracrine, or endocrine effects and include IGF-1 and IGF-2 molecules, which are involved in enhancing vascular endothelial activity in ischaemic vessels (31). IGF-1 and IGF-2 are found in various tissues, including muscle tissue, which acts in an endocrine way by targeting other organs including the kidneys, and therefore, could carry out a crosstalk between muscle and bone cells (32).
Indeed, it seems that IGF-1 does not protect against the pro-inflammatory and apoptotic action induced by cytokines. A protective effect occurs with IGF-2, which by binding to the IGF-1R receptor, induces IL-10, an anti-inflammatory cytokine which protects islet transplantation and improves survival (18).
In rheumatic diseases, the severity of the pathological state may depend on the balance between pro-inflammatory and anti-inflammatory cytokines. In rheumatoid arthritis (RA), the joint-inflamed site harbors many pro-inflammatory cytokines that belong to the IL-1 family such as IL-1, IL-18, IL-33, IL-36α, IL-36β and IL-36γ; but anti-inflammatory anti-receptors can also be found, such as the IL-1 receptor antagonist (IL-1Ra) and IL-36 receptor antagonist (IL-36Ra), and two cytokines, IL-37 and IL- 38, which inhibit innate immunity and inflammation. IL-37 acts as a suppressor by inhibiting mammalian target of rapamycin (mTOR) and increases AMP kinase activity. IL-38 performs its anti-inflammatory activity by binding to the IL-1R6 receptor, a complex that causes the recruitment of IL-1R9.
IGF-2 regulates bone development by acting on chondrocytes, osteoblasts, osteocytes, and osteoclasts, and is implicated in skeletal ageing (33). In fact, the serum levels of IGF-2 tend to decrease with the ageing of the bones, an action that takes place with the reduction of the mineral bone density (33).
IGFBP is involved in many biological activities including cartilage failure during osteoarthritis (OA) (34). In the synovial fluid and cartilage of OA patients, it was seen that IGFBP levels were increased (35), an effect that implicates IGF-1 and 2 as important factors in inflammation. IGFBP overexpression could influence the availability of IGFs with alteration of the chondrocyte vital pathway.
Chondrocyte hypertrophy is characterized by an increase in oxidative phosphorylation which is regulated by IGF-2. In a recent interesting article, Hollander JM et al., reported that the chondrocyte maturation during cartilage development indicates that the inhibition of oxidative phosphorylation in murine chondrocytes can damage hypertrophic differentiation (36). These authors reported that an IGF-2 deficiency can result in increased oxidative phosphorylation in hypertrophic chondrocytes. The results demonstrate that IGF-2 is important in evading excessive glucose metabolism and is determinant for bone development (36).
CONCLUSIONS
In conclusion, in this study we highlighted that the inflammatory cytokines IL-1 and TNF mediate the immune response with the participation of IGFs. During inflammation, there are also neuroendocrine and metabolic responses with pituitary generation of IGF. Furthermore, IGF-1 and IGF-2 have different biological effects. IGF-1 mediates bone homeostasis and activates the MAPK pathway, resulting in cell proliferation. IGF-2 plays a crucial role in the physiological development and differentiation of the mouse embryo mesoderm, favoring tumor growth and influencing cellular proliferative pathologies. A protective effect of IGF-2 is induced by the activation of IL-10, while IGF-1 does not protect against pro-inflammatory and apoptotic responses. Moreover, IGF-2 mediates chondrocyte hypertrophy with an increase in oxidative phosphorylation.
Conflict of interest
The authors declare that they have no conflict of interest.
REFERENCES
- Pérez S, Rius-Pérez S. Macrophage Polarization and Reprogramming in Acute Inflammation: A Redox Perspective. Antioxidants. 2022;11(7):1394. doi:https://doi.org/10.3390/antiox11071394
- Li Z, Xiao Fang Yu, Werner J, Bazhin AV, D’Haese JG. The role of interleukin-18 in pancreatitis and pancreatic cancer. Cytokine Growth Factor Rev. 2019;50:1-12. doi:https://doi.org/10.1016/j.cytogfr.2019.11.001
- Hu F, Lou N, Jiao J, Guo F, Xiang H, Shang D. Macrophages in pancreatitis: Mechanisms and therapeutic potential. Biomedicine & Pharmacotherapy. 2020;131:110693. doi:https://doi.org/10.1016/j.biopha.2020.110693
- Ren Y, Zhao H, Yin C, et al. Adipokines, Hepatokines and Myokines: Focus on Their Role and Molecular Mechanisms in Adipose Tissue Inflammation. Frontiers in Endocrinology. 2022;13:873699. doi:https://doi.org/10.3389/fendo.2022.873699
- Baxter RC. Insulin-like growth factor (IGF)-binding proteins: interactions with IGFs and intrinsic bioactivities. American Journal of Physiology Endocrinology and Metabolism. 2000;278(6):E967-976. doi:https://doi.org/10.1152/ajpendo.2000.278.6.E967
- Frystyk J, Skjaerbaek C, Dinesen B, Ørskov H. Free insulin-like growth factors (IGF-I and IGF-II) in human serum. FEBS Letters. 1994;348(2):185-191. doi:https://doi.org/10.1016/0014-5793(94)00602-4
- Gomarasca M, Banfi G, Lombardi G. Myokines: The endocrine coupling of skeletal muscle and bone. Advances in Clinical Chemistry. 2020;94:155-218. doi:https://doi.org/10.1016/bs.acc.2019.07.010
- Jagodzik P, Tajdel-Zielinska M, Ciesla A, Marczak M, Ludwikow A. Mitogen-Activated Protein Kinase Cascades in Plant Hormone Signaling. Frontiers in Plant Science. 2018;9. doi:https://doi.org/10.3389/fpls.2018.01387
- Niu T, Rosen CJ. The insulin-like growth factor-I gene and osteoporosis: A critical appraisal. Gene. 2005;361:38-56. doi:https://doi.org/10.1016/j.gene.2005.07.016
- Guntur AR, Rosen CJ. IGF-1 regulation of key signaling pathways in bone. BoneKEy Reports. 2013;2. doi:https://doi.org/10.1038/bonekey.2013.171
- Yakar S, Adamo ML. Insulin-Like Growth Factor-1 Physiology: Lessons from Mouse Models. Endocrinology and metabolism clinics of North America. 2012;41(2):231-v. doi:https://doi.org/10.1016/j.ecl.2012.04.008
- Bikle DD, Tahimic C, Chang W, Wang Y, Philippou A, Barton ER. Role of IGF-I signaling in muscle bone interactions. Bone. 2015;80:79-88. doi:https://doi.org/10.1016/j.bone.2015.04.036
- Han VKM, D’Ercole AJ, Lund PK. Cellular Localization of Somatomedin (Insulin-Like Growth Factor) Messenger RNA in the Human Fetus. Science. 1987;236(4798):193-197. doi:https://doi.org/10.1126/science.3563497
- Firth SM, Baxter RC. Cellular Actions of the Insulin-Like Growth Factor Binding Proteins. Endocrine Reviews. 2002;23(6):824-854. doi:https://doi.org/10.1210/er.2001-0033
- DeChiara TM, Efstratiadis A, Robertsen EJ. A growth-deficiency phenotype in heterozygous mice carrying an insulin-like growth factor II gene disrupted by targeting. Nature. 1990;345(6270):78-80. doi:https://doi.org/10.1038/345078a0
- Rotwein P, Pollock KM, Watson M, Milbrandt JD. Insulin-Like Growth Factor Gene Expression during Rat Embryonic Development*. Endocrinology. 1987;121(6):2141-2144. doi:https://doi.org/10.1210/endo-121-6-2141
- Lui JC, Finkielstain GP, Barnes KM, Baron J. An imprinted gene network that controls mammalian somatic growth is down-regulated during postnatal growth deceleration in multiple organs. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 2008;295(1):R189-R196. doi:https://doi.org/10.1152/ajpregu.00182.2008
- Belfiore A, Rosaria Valentina Rapicavoli, Rosario Le Moli, et al. IGF2: A Role in Metastasis and Tumor Evasion from Immune Surveillance? Biomedicines. 2023;11(1):229-229. doi:https://doi.org/10.3390/biomedicines11010229
- Wang X, Lin L, Lan B, et al. IGF2R-initiated proton rechanneling dictates an anti-inflammatory property in macrophages. Sci Adv. 2020;6(48). doi:https://doi.org/10.1126/sciadv.abb7389
- Wise TL, Pravtcheva DD. Delayed Onset of Igf2-Induced Mammary Tumors in Igf2r Transgenic Mice. Cancer Res. 2006;66(3):1327-1336. doi:https://doi.org/10.1158/0008-5472.can-05-3107
- Malecova B, Gatto S, Etxaniz U, et al. Dynamics of cellular states of fibro-adipogenic progenitors during myogenesis and muscular dystrophy. Nature Communications. 2018;9(1):3670. doi:https://doi.org/10.1038/s41467-018-06068-6
- Forcina L, Miano C, Scicchitano BM, Musarò A. Signals from the Niche: Insights into the Role of IGF-1 and IL-6 in Modulating Skeletal Muscle Fibrosis. Cells. 2019;8(3). doi:https://doi.org/10.3390/cells8030232
- Torrente Y, Bella P, Tripodi L, Villa C, Farini A. Role of Insulin-Like Growth Factor Receptor 2 across Muscle Homeostasis: Implications for Treating Muscular Dystrophy. Cells. 2020;9(2):441-441. doi:https://doi.org/10.3390/cells9020441
- Maggio M, De Vita F, Lauretani F, et al. IGF-1, the cross road of the nutritional, inflammatory and hormonal pathways to frailty. Nutrients. 2013;5(10):4184-4205. doi:https://doi.org/10.3390/nu5104184
- Straub RH, Cutolo M, Buttgereit F, Pongratz G. Review: Energy regulation and neuroendocrine-immune control in chronic inflammatory diseases. Journal of Internal Medicine. 2010;267(6):543-560. doi:https://doi.org/10.1111/j.1365-2796.2010.02218.x
- Thomas FH, EthierJF, Shimasaki S, Vanderhyden BC. Follicle-Stimulating Hormone Regulates Oocyte Growth by Modulation of Expression of Oocyte and Granulosa Cell Factors. Endocrinology. 2005;146(2):941-949. doi:https://doi.org/10.1210/en.2004-0826
- Rama Raju G, Chavan R, Deenadayal M, et al. Luteinizing hormone and follicle stimulating hormone synergy: A review of role in controlled ovarian hyper-stimulation. Journal of Human Reproductive Sciences. 2013;6(4):227.
- Agis-Balboa RC, Arcos-Diaz D, Wittnam J, et al. A hippocampal insulin-growth factor 2 pathway regulates the extinction of fear memories. The EMBO Journal. 2011;30(19):4071-4083. doi:https://doi.org/10.1038/emboj.2011.293
- Hughes A, Rojas-Canales D, Drogemuller C, Voelcker NH, Grey ST, Coates PTH. IGF2: an endocrine hormone to improve islet transplant survival. Journal of Endocrinology. 2014;221(2):R41-R48. doi:https://doi.org/10.1530/joe-13-0557
- Severinsen MCK, Pedersen BK. Muscle–Organ Crosstalk: The Emerging Roles of Myokines. Endocrine Reviews. 2020;41(4):594-609. doi:https://doi.org/10.1210/endrev/bnaa016
- Higashi Y, Quevedo HC, Tiwari S, et al. The Interaction Between IGF-1, Atherosclerosis and Vascular Aging. Frontiers of hormone research. 2014;43:107. doi:https://doi.org/10.1159/000360571
- Pedersen BK, Febbraio MA. Muscles, exercise and obesity: skeletal muscle as a secretory organ. Nature reviews Endocrinology. 2012;8(8):457-465. doi:https://doi.org/10.1038/nrendo.2012.49
- Yakar S, Werner H, Rosen CJ. 40 YEARS OF IGF1: Insulin-like growth factors: actions on the skeleton. Journal of Molecular Endocrinology. 2018;61(1):T115-T137. doi:https://doi.org/10.1530/jme-17-0298
- Galasso O, M De Gori, Nocera A, Brunetti A, Gasparini G. Regulatory Functions of Insulin-like Growth Factor Binding Proteins in Osteoarthritis. Int J Immunopathol Pharmacol. 2011;24(1_suppl2):55-59. doi:https://doi.org/10.1177/03946320110241s211
- Matsumoto T, Gargosky SE, Iwasaki K, Rosenfeld RG. Identification and characterization of insulin-like growth factors (IGFs), IGF-binding proteins (IGFBPs), and IGFBP proteases in human synovial fluid. J Clin Endocrinol Metab. 1996;81(1):150-155. doi:https://doi.org/10.1210/jcem.81.1.8550744
- Hollander JM, Li L, Rawal M, et al. A critical bioenergetic switch is regulated by IGF2 during murine cartilage development. Commun Biol. 2022;5(1). doi:https://doi.org/10.1038/s42003-022-04156-4